График работы:
E-mail:
Бесплатная консультация
((R,S)-2-(2-оксо-4-фенилпирролидин-1-ил)ацетамид, карфендон, фенотропил), более известный как фенилпирацетам. Фенилпирацетам представляет собой рацемическую смесь как R, так и S энантиомеров и классифицируется как ноотропное соединение, которое в настоящее время изучается на предмет его потенциала в борьбе с когнитивным снижением, ожирением, усталостью и депрессией. Недавние исследования показали, что R-фенилпирацетам отвечает за стимуляцию локомоции соединения, а также работает совместно с S-фенилпирацетамом для ингибирования обратного захвата дофамина в синаптической щели. Кроме того, было показано, что R-фенилпирацетам связывается с транспортером норадреналина (NET) и, как правило, более активен, чем энантиомер S-фенилпирацетама [1].
1) Селективные ингибиторы дофамина, такие как S-фенилпирацетам, могут иметь потенциал для лечения ожирения у лиц с диагнозом метаболический синдром с меньшим количеством случаев возникновения неблагоприятных побочных эффектов и последствий для здоровья.
2) Было показано, что R-фенилпирацетам обладает потенциальным нейропротекторным и противовоспалительным действием, что указывает на то, что соединения, ингибирующие активность транспортеров дофамина, могут использоваться в качестве эффективного лечения когнитивных нарушений.
1) Исследовательская группа Звейниеце и др. изучала влияние S-фенилпирацетама на набор веса, уровни глюкозы и лептина, а также общую двигательную активность в экспериментальной модели на животных. Набор веса обычно вызван избыточным потреблением энергии и характеризуется увеличением жировой ткани, что приводит к более высокому риску сопутствующих заболеваний. Предыдущие исследования показали, что дофаминергические пути играют решающую роль в контроле веса тела и потребления пищи, учитывая, что визуализация указывает на сниженную доступность дофаминовых рецепторов у людей с ожирением по сравнению с людьми со здоровым ИМТ [1].
Для части исследования in vivo шесть самцов крыс Wistar были декапитированы для того, чтобы препарировать мозг, чтобы исследователи могли изолировать полосатое тело. После выделения полосатое тело гомогенизировали в буфере сахарозы-фосфата с последующим центрифугированием в течение 10 минут. Полученный супернатант снова центрифугировали, а осадок, полученный в результате этого конечного процесса, ресуспендировали в буфере натрия-фосфата. Анализ связывания проводился с использованием образцов, состоящих из 60 мкл буфера натрия-фосфата, 20 мкл фенилпирацетама, 20 мкл воды, 100 мкл аликвот мембран и 20 мкл [3H}WIN 35,428 [1]. Анализы неспецифического связывания проводились с использованием тех же образцов с добавлением дигидрохлорида ваноксерина с последующей инкубацией на льду в течение 2 часов. Наконец, для измерения радиоактивности, как свободные, так и связанные радиолиганды были разделены с помощью быстрой фильтрации, достигнутой путем трехкратной промывки фильтров ледяным буфером TRIS-HCl. Этот процесс позволил измерить радиоактивность методом жидкостной сцинтилляции.
Для части исследования in vitro использовались шестнадцать 6-недельных тучных самцов крыс Zucker весом от 130 до 200 граммов. Этих испытуемых сравнивали с восемью крысами Zucker того же возраста, которые были более худыми и весили от 130 до 170 граммов. Крысам давали возможность адаптироваться к их местным условиям в течение 7 дней до начала лечения; испытуемые содержались в стандартных лабораторных условиях с неограниченным доступом к пище и воде. Тучным крысам Zucker ежедневно вводили дозу S-фенилпирацетама 50 мг/кг через пероральный зонд в течение 12 недель. Худым крысам Zucker вводили то же самое, однако вместо S-фенилпирацетама им вводили 50 мг/кг воды [1].
Следующая часть эксперимента in vitro включала 32 самца мышей C57BL/6N весом от 21 до 25 граммов. Мышам давали возможность адаптироваться к местным условиям в течение 7 дней до начала экспериментов; испытуемые содержались в стандартных лабораторных условиях с неограниченным доступом к пище и воде. После периода адаптации испытуемые были случайным образом распределены по четырем различным группам лечения, включая: нормальную диету, западную диету, западную диету с лечением S-фенилпирацетамом и западную диету с лечением R-фенилпирацетамом. Важно отметить, что западная диета была охарактеризована как диета, содержащая 21% жира, 20% белка и 50% углеводов, в то время как нормальная диета содержала 4,5% жира, 14,5% белка и 60,1% углеводов [1]. Мышам, получавшим активное лечение R- или S-фенилпирацетамом, вводили 50 мг/кг через желудочный зонд, в то время как мышам, получавшим как западную диету, так и обычную диету, вводили 50 мг/кг воды тем же способом.
Последняя экспериментальная группа in vitro включала сорок самцов мышей SW в возрасте 16 недель весом от 40 до 50 граммов каждый. Испытуемые содержались в стандартных лабораторных условиях с неограниченным доступом к пище и воде. Мыши были случайным образом распределены по пяти различным группам, включая: введение 50 мг/кг R-фенилпирацетама, введение 100 мг/кг R-фенилпирацетама, введение 50 мг/кг S-фенилпирацетама, введение 100 мг/кг S-фенилпирацетама и введение питьевой воды для контрольной группы [1]. Все животные, включенные в эксперимент, взвешивались один раз в неделю, чтобы сделать выводы относительно увеличения жировой массы путем сравнения различий между массой животных с ожирением и ожирением. Конкретная формула для расчета жировой массы, особенно у субъектов на западной диете, была следующей: жировая масса = средний вес животного с ожирением – средний вес животного с ожирением. После 12 недель лечения и изменения диеты испытуемые были помещены в индивидуальные метаболические клетки для измерения потребления ими пищи.
Затем оценивали уровни толерантности к глюкозе и концентрации в крови. Перед тестированием толерантности к глюкозе мышей и крыс выдерживали на голодной диете в течение ночи, после чего им внутрибрюшинно вводили раствор глюкозы в дозах 1 г/кг для крыс и 0,5 г/кг для мышей. Образцы крови брали из хвостовой вены через 15, 30, 60, 120, 180 и 240 минут после введения раствора глюкозы. Тест на толерантность к глюкозе проводили через 8 недель лечения мышей и через 12 недель лечения S-фенилпирацетамом для крыс. Те же временные рамки использовались при измерении концентрации глюкозы в плазме крови, а также при измерении концентраций лептина и инсулина в плазме. Значения концентрации глюкозы оценивали с помощью коммерчески доступного набора от Instrumentation Laboratory, в то время как лептин и инсулин оценивали с помощью наборов ELISA для мышиного лептина и крысиного/мышиного инсулина [1].
Наконец, испытуемые прошли тест открытого поля, чтобы наблюдать изменения двигательной активности у животных после 8 недель лечения S-фенилпирацетамом для мышей и 12 недель лечения для крыс. Аппарат, используемый в тесте, представлял собой квадратную арену, в центре которой размещались животные, чтобы исследовательская группа могла зафиксировать пройденное расстояние в см и скорость движения в см/секунду с помощью системы видеослежения. Тест открытого поля проводился через 60 минут после введения энантиомеров фенилпирацетама; мышей помещали в открытое поле на 4-минутный период тестирования, а крыс помещали в открытое поле на 15-минутный период тестирования.
В качестве дополнительного испытания исследовательская группа использовала тест открытого поля для изучения двигательной активности крыс и мышей после однократного введения S- и R-фенилпирацетама, а не после 8-12 недель лечения. Испытуемым разрешалось исследовать аппарат в течение 10 минут за день до эксперимента, после чего они возвращались в свои исходные клетки. Через 24 часа после периода адаптации эксперимент проводился в два сеанса по 12 минут, которые проводились через 30 минут после введения S-фенилпирацетама и через 60 минут после введения R-фенилпирацетама. Животных помещали обратно в центр открытого поля, чтобы можно было записать скорость и пройденное расстояние [1].
2) В то время как ранее упомянутое исследование было сосредоточено на эффективности S-фенилпирацетама, исследовательская группа Звейниеце и др. теперь изучила, как R-фенилпирацетам изменил противовоспалительную активность транспорта дофамина в воспалительных моделях животных исследования. Для целей этого эксперимента исследователи использовали 246 мышей ICR в возрасте 8-10 недель, каждая весом 23-25 граммов. Животных по отдельности размещали в вентилируемой клетке и содержали в стандартных лабораторных условиях, и им был предоставлен свободный доступ к пище и воде. Оттуда мышей случайным образом распределяли по различным группам лечения для начала эксперимента и получали как внутрибрюшинную, так и пероральную дозу 50 мг/кг за 15 минут, 30 минут, 1, 2, 4, 6 и 24 часа до того, как субъекты были подвергнуты эвтаназии с целью сбора образцов мозговой ткани [2].
Мышам также делали инъекцию каррагинана интраплантарно, чтобы вызвать отек лапы мыши. Перед инъекцией каррагинана всем испытуемым перорально вводили либо 10 мг/кг физиологического раствора, либо 25 или 50 мг/кг R-фенилпирацетама, либо 10 мг/кг индометацина в течение экспериментального периода продолжительностью 7 дней. Важно отметить, что вторичная часть теста с каррагинаном сравнивала как R, так и S энантиомеры путем введения пероральной дозы 50 мг/кг каждого в течение экспериментального периода продолжительностью 7 дней. В последний день лечения животные получали свою лечебную дозу за 60 минут до инъекции каррагинана интраплантарно в правую лапу; половине испытуемых вводили 40 мкл физиологического раствора или 40 мкл 2% каррагинана. После инъекции исследовательская группа отслеживала изменения в процентном отношении объема лапы, связанного с вызванным отеком [2].
В связи с тестом с каррагинаном испытуемый также прошел электронный тест фон Фрея, чтобы исследовать механическую аллодинию через порог отдергивания лапы, в которую был введен каррагинан. Через 7 часов после инъекции был использован электронный анестезиометр фон Фрея для применения механического стимула. Все испытуемые были помещены на сетчатый пол в индивидуальных камерах наблюдения за 10 минут до испытания, чтобы акклиматизироваться к окружающей среде. Период адаптации немедленно сопровождался размещением нити фон Фрея на срединно-подошвенной поверхности правой задней лапы, в то время как исследователи регистрировали силу, необходимую для отдергивания лапы.
Исследовательская группа также подвергла мышей тесту на облизывание лап, вызванному формалином. Перед тестированием контрольные животные получили введение физиологического раствора, в то время как животные в экспериментальных группах получили лечение либо 10, либо 50 мг/кг R-фенилпирацетама, либо лечение 50 мг/кг S-фенилпирацетама в течение экспериментального периода в 7 дней. В последний день лечения животным вводили их последнюю дозу за 60 минут до того, как им интрапланарно вводили 30 мкл раствора формалина в правую заднюю лапу. Затем каждого испытуемого помещали в отдельные клетки для наблюдения, чтобы исследовательская группа могла записать общее время облизывания задней лапы каждой мыши в течение трех отдельных фаз продолжительностью 5-10 минут каждая [2].
За шесть часов до эвтаназии и взятия образцов мозговой ткани мышам делали внутрибрюшинную инъекцию 20 мг/кг ЛПС для получения данных относительно экспрессии воспалительных генов. ЛПС вводили одновременно с дозой 50 мг/кг R-фенилпирацетама или S-фенилпирацетама у экспериментальных животных или с инъекцией физиологического раствора у контрольных животных [2]. ЛПС и лечебные соединения вводили в контралатеральные стороны, а ректальную температуру испытуемых измеряли как до инъекции, так и через 6 часов после, перед декапитацией и сбором образцов мозга и крови.
Образцы крови, собранные у мышей, были пропущены через центрифугирование в течение 10 минут, чтобы изолировать плазму. Ткань мозга была разделена на правое и левое полушария с последующей гомогенизацией, полученный гомогенат снова центрифугировали в течение еще 10 минут, чтобы оставить супернатант, который был гомогенизирован и центрифугирован в последний раз. Полученный осадок был заморожен до дальнейшего анализа. Образцы изолированной плазмы и конечного супернатанта были приготовлены в процессе депротеинизации с последующим объединением 100 мкл образца и 500 мкл 0,1% раствора муравьиной кислоты в ацетонитриле. Подготовленные и депротеинизированные образцы центрифугировали в течение 20 минут и переносили в экспериментальные флаконы для сверхэффективной жидкостной хроматографии-тандемной масс-спектрометрии [2].
1) При рассмотрении анализа связывания, связанного с транспортировкой дофамина после лечения R-фенилпирацетамом и S-фенилпирацетамом, результаты показали, что оба энантиомера ноотропа конкурентно связывались с участками транспорта дофамина, успешно ингибируя обратный захват дофамина в синаптической щели. В дополнение к транспортировке дофамина, эффекты, вызванные S-фенилпирацетамом на локомоторную активность, наблюдались в ходе тестирования в открытом поле, которое проводилось через 30 минут после введения ноотропа. Было показано, что лечение S-энантиомером фенилпирацетама значительно увеличивает локомоторную активность в течение первых 4 минут 12-минутного испытания. С 4-8 и 8-12 минут локомоция непрерывно снижалась. Важно отметить, что доза 100 мг/кг R-фенилпирацетама, как было показано, увеличивает локомоторную активность в большей степени, чем та же доза S-фенилпирацетама [1].
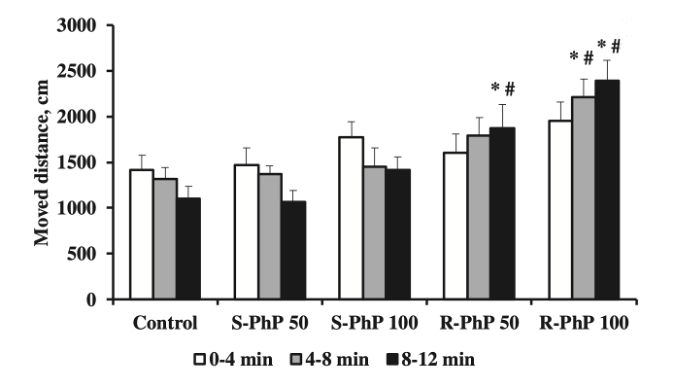
Рисунок 1: Зарегистрированные уровни двигательной активности через 30 минут после введения R-фенилпирацетама или S-фенилпирацетама.
При изучении эффектов S-фенилпирацетама на набор веса исследователь использовал двухфакторный дисперсионный анализ (ANOVA), который выявил значительную корреляцию между днем лечения и экспериментальной группой, в которую были включены крысы Цукера. Результаты показали, что по сравнению с контрольными крысами с худым весом контрольные крысы с ожирением испытали резкое увеличение веса тела, начиная со 2-й недели, которое сохранялось до конца эксперимента. Увеличение жировой массы было измерено и составило 5 граммов после первой недели исследования; это измерение продолжало увеличиваться в течение оставшегося периода экспериментального времени и было зафиксировано как 173 грамма к концу 12 недель.
У тучных крыс, получавших S-фенилпирацетам, скорость набора веса тела была значительно снижена по сравнению с контрольной группой с ожирением. Этот результат наблюдался, начиная с 4-й недели эксперимента и сохранялся до конца эксперимента. После 12 недель лечения ноотропом исследователи зафиксировали 16%-ное снижение веса тела. Что касается потребления пищи, не было выявлено существенных различий между контрольными крысами с ожирением и худыми, однако группы, получавшие S-фенилпирацетам, продемонстрировали тенденцию к потреблению меньшего количества пищи по сравнению с контрольными крысами с ожирением [1].
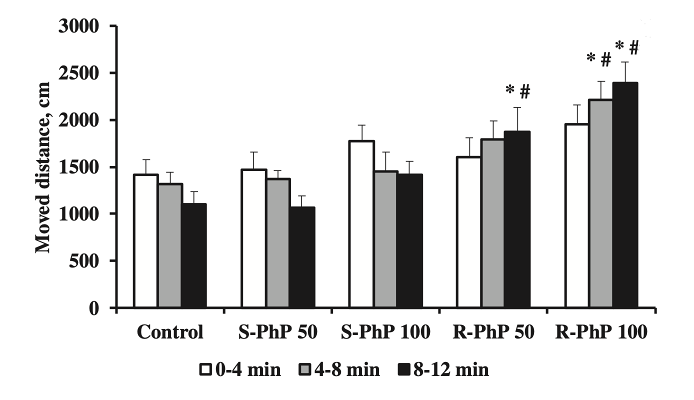
Рисунок 2: Изменения в A) увеличении массы тела, B) увеличении жировой массы и C) потреблении пищи в контрольных группах худых и тучных мышей, а также в экспериментальной группе мышей с ожирением, которым вводили S-фенилпирацетам.
Начиная со 2-й недели эксперимента, было обнаружено, что прирост массы тела был значительно выше у мышей, питавшихся по западной диете, чем у контрольных мышей, питавшихся по обычной диете. Как уже упоминалось ранее, двухфакторный дисперсионный анализ выявил корреляцию между днем лечения и экспериментальной группой. Соответствующие результаты эксперимента показали, что лечение R-фенилпирацетамом вызвало наиболее значительное снижение массы тела в течение первых двух недель исследования, а также на 6-8-й неделе. Было обнаружено, что лечение S-фенилпирацетамом вызвало наиболее значительное снижение массы тела, начиная с 6-й недели и сохраняясь до конца 12-недельного экспериментального периода.
К 8 неделе группа, получавшая S-фенилпирацетам, испытала 26% снижение массы тела, в то время как среднее снижение массы тела в группе, получавшей R-фенилпирацетам, составило 40%. Кроме того, наблюдалось значительное увеличение жировой массы у мышей, получавших западную диету, по сравнению с мышами, получавшими обычную диету. Однако мыши, получавшие S-фенилпирацетам, испытали 45% снижение жировой массы к 8-й неделе лечения, в то время как мыши, получавшие R-фенилпирацетам, испытали 69% снижение жировой массы по сравнению с контрольной группой, получавшей западную диету [1].
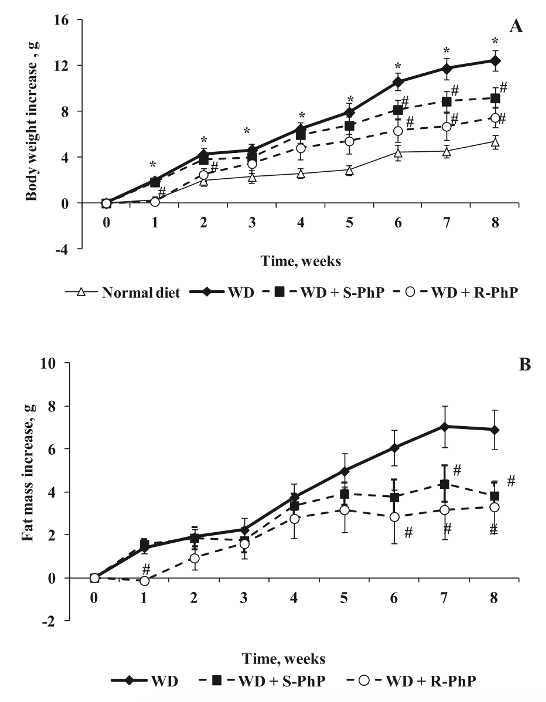
Рисунок 3: Зависящие от времени изменения A) массы тела и B) жировой массы у мышей, получавших обычную диету, западную диету, западную диету с введением S-фенилпирацетама и западную диету с введением R-фенилпирацетама.
Наконец, исследовательская группа изучила изменения уровней глюкозы в крови, лептина и инсулина в ответ на лечение как S-, так и R-фенилпирацетамом. Исходные уровни у контрольных крыс с ожирением и худых были схожими; контрольные крысы с ожирением испытали увеличение концентрации глюкозы в крови натощак, начиная с 8 недель после начала эксперимента. Через 12 недель как голодные, так и накормленные контрольные крысы с ожирением испытывали гипергликемию, однако у субъектов, получавших лечение S-фенилпирацетамом, была значительно более низкая концентрация глюкозы в крови в состоянии сытости по сравнению с контрольной группой с ожирением. Следует отметить, что лечение ноотропом не вызвало никаких изменений в концентрации глюкозы в крови у голодных мышей. При изучении данных, полученных в ходе теста на толерантность к глюкозе, исследователи смогли сделать вывод, что введение S-фенилпирацетама улучшило толерантность к глюкозе по сравнению с контрольными мышами с ожирением [1].
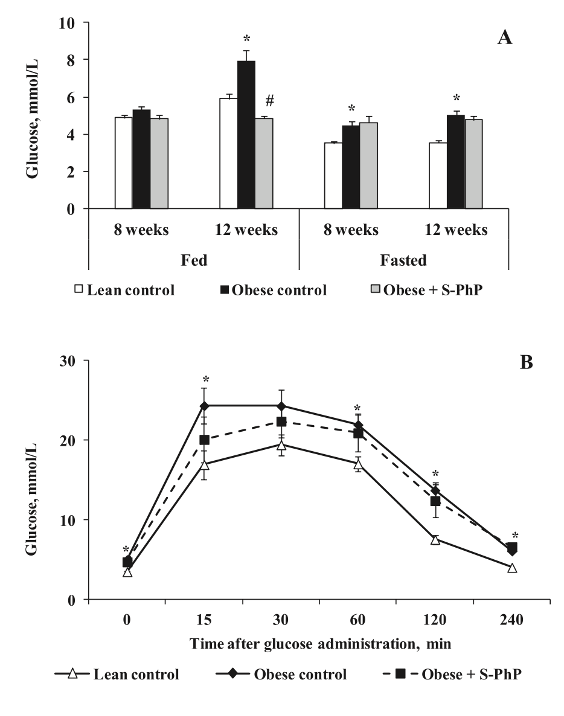
Рисунок 4: Изменения A) уровня глюкозы в крови и B) толерантности к глюкозе у контрольных мышей с ожирением и худых мышей и у мышей с ожирением, получавших S-фенилпирацетам
2) Результаты исследования, проведенного Звейниеце и др., сначала были сосредоточены на результатах, наблюдаемых после инъекции ЛПС для индукции гипотермии и сверхэкспрессии генов. Первоначальные результаты показали, что наблюдалось значительное снижение температуры тела у контрольных животных, когда температура измерялась через 6 часов после инъекции ЛПС. При этом предварительная обработка как R-, так и S-энантиомерами была способна восстановить гипотермию, вызванную ЛПС. Кроме того, было показано, что инъекция ЛПС увеличивает экспрессию гена IL-1 бета в 8 раз, TNF-альфа в 19 раз и iNOS в 12 раз. Предварительная обработка R-фенилпирацетамом снизила экспрессию IL-1 бета на 75%, TNF-альфа на 73% и iNOS на 65% [2]. С другой стороны, предварительное лечение S-фенилпирацетамом лишь немного снизило экспрессию генов ИЛ-1 бета и ФНО-альфа, однако важно отметить, что наблюдалась значительная разница между экспрессией iNOS у пациентов, получавших лечение R-фенилпирацетамом, по сравнению с пациентами, получавшими лечение S-фенилпирацетамом.
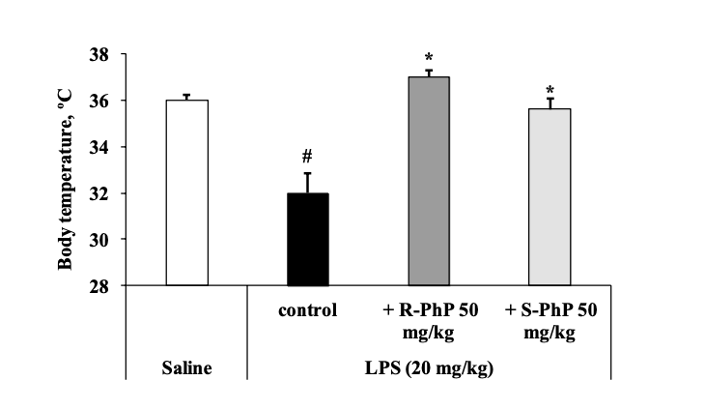
Рисунок 5: Изменения температуры тела в ответ на предварительную обработку ноотропными энантиомерами после инъекции ЛПС
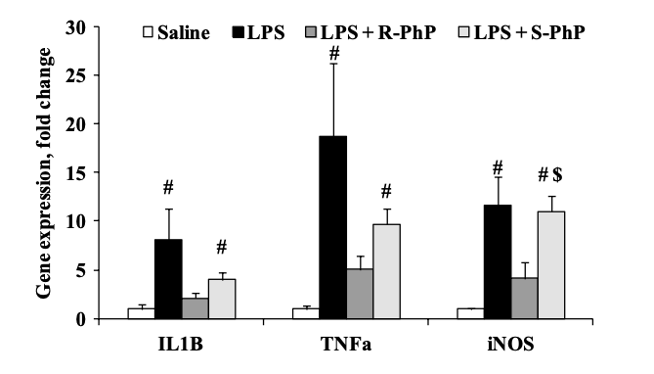
Рисунок 6: Изменения в экспрессии генов ИЛ-1 бета, ФНО-альфа и iNOS в ответ на предварительную обработку ноотропными энантиомерами после инъекции ЛПС.
Каррагинан вводили в правую заднюю лапу мышей, чтобы оценить потенциал фенилпирацетама для усиления воспаления, измеряемого по степени отека лапы. Результаты этой части исследования показали, что каррагинан увеличивал отек лапы в зависимости от дозы у мышей, которым вводили физиологический раствор, а не экспериментальную дозировку. Было обнаружено, что предварительная обработка 50 мг/кг R-фенилпирацетама уменьшала отек в зависимости от времени. Через 2 часа после инъекции отек уменьшился на 60%, через 4 часа после инъекции отек уменьшился на 40% и через 6 часов после инъекции отек уменьшился на 45% [2]. Более низкие дозы R-фенилпирацетама не вызвали значительного уменьшения объема лапы, однако дозы индометацина 10 мг/кг значительно уменьшили отек лапы на 36% на 4-часовой отметке и на 45% на 6-часовой отметке после инъекции каррагинана. Связанные с инъекцией каррагинана результаты электронного теста с нитью фон Фрея показали, что время, необходимое для наступления отстранения, сократилось на 35%, когда мышам предварительно вводили 50 мг/кг R-фенилпирацетама. Кроме того, время, необходимое для наступления отстранения, сократилось на 40%, когда испытуемым вводили 10 мг/кг индометацина [2].
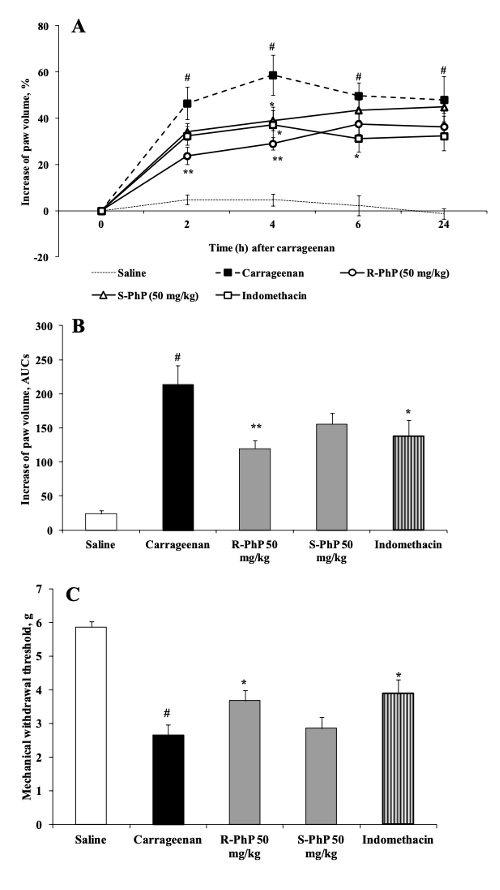
Рисунок 7: Изменения объема лапы (A и B) и порога механического отдергивания (C) в ответ на введение R-фенилпирацетама и S-фенилпирацетама
Наконец, эффекты ингибирования боли ноотропными энантиомерами были исследованы с помощью теста на облизывание лап, вызванного формалином. Контрольные мыши, которым вводили физиологический раствор, испытывали немедленную ноцицептивную реакцию после инъекции формалина. Ноцицептивная реакция включала встряхивание и облизывание инъецированной задней лапы, которые продолжались на протяжении всех трех фаз эксперимента. Было обнаружено, что лечение R-фенилпирацетамом в дозах 10 и 50 мг/кг сокращает продолжительность облизывания лапы в зависимости от дозы; было показано, что доза 50 мг/кг сокращает облизывание лапы на 50% во время второй фазы [2].
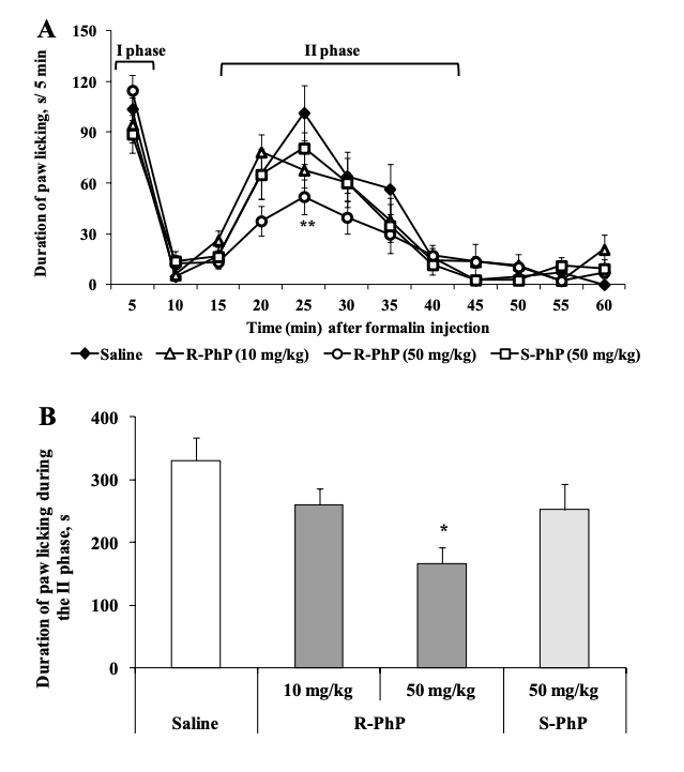
Рисунок 8: Изменения продолжительности облизывания лап.
[1] Zvejniece L, Svalbe B, Vavers E, Makrecka-Kuka M, Makarova E, Liepins V, Kalvinsh I, Liepinsh E, Dambrova M. S-phenylpiracetam, a selective DAT inhibitor, reduces body weight gain without influencing locomotor activity. Pharmacol Biochem Behav. 2017 Sep;160:21-29. doi: 10.1016/j.pbb.2017.07.009. Epub 2017 Jul 22. PMID: 28743458.
[2] Zvejniece L, Zvejniece B, Videja M, Stelfa G, Vavers E, Grinberga S, Svalbe B, Dambrova M. Neuroprotective and anti-inflammatory activity of DAT inhibitor R-phenylpiracetam in experimental models of inflammation in male mice. Inflammopharmacology. 2020 Oct;28(5):1283-1292. doi: 10.1007/s10787-020-00705-7. Epub 2020 Apr 11. PMID: 32279140.
© 2026 АнкеБио. Magazin nootropov Biohaker
Казахстан, Алматы, ул. Байзакова, 280
Все права защищены.
+7 (977) 520-97-00
Email: ankebiorus@gmail.com
Вы нашли для себя препарат, который хотите
приобрести по низкой цене и с гарантией
безопасной доставки?
Звоните нам
Мы Вам обязательно ответим!