
График работы:
E-mail:
Бесплатная консультация
Нефирацетам классифицируется как ноотропное средство, которое является производным пирролидона. Недавно проведенное исследование на животных показало, что это соединение облегчает когнитивную функцию и уменьшает амнезию, вызванную лекарственными средствами. Предполагаемый механизм действия нефирацетама связан со способностью производных пирролидона активировать дофаминергические, холинергические и глутаматергические нейротрансмиттерные системы. Активация этих систем приводит к увеличению высвобождения нейротрансмиттеров из пресинаптических окончаний и улучшению высвобождения нейротрансмиттеров из постсинаптических окончаний. Текущие исследования, касающиеся ноотропа, сосредоточены на его способности лечить когнитивные нарушения, связанные с травматическими повреждениями и нейродегенерацией [1].
1) На основании результатов теста с водным лабиринтом Морриса, лечение нефирацетамом может эффективно лечить когнитивную дисфункцию, связанную с черепно-мозговой травмой.
2) При оценке классического условного рефлекса моргания как показателя поведенческой парадигмы и когнитивного функционирования у кроликов было показано, что введение нефирацетама смягчает дефицит обучения у старых животных.
1) Эксперимент, проведенный исследовательской группой ДеФорда и соавторов, изучал способность нефирацетама способствовать восстановлению когнитивных функций после черепно-мозговой травмы, вызванной у испытуемых. Для целей данного исследования для тестирования были использованы 39 взрослых самцов крыс Sprague-Dawley весом 300-350 граммов. Крысы содержались в стандартных условиях с неограниченным доступом к пище и воде. Все испытуемые были случайным образом распределены на 5 различных групп, включая: нелеченная ложная травма, ложная травма + 9 мг/кг нефирацетама, травма-физиологический раствор, травма + 3 мг/кг нефирацетама, травма + 9 мг/кг нефирацетама. Ноотроп вводили животным перорально между 9:00 и 11:00 утра с 1-го по 15-й день после индукции черепно-мозговой травмы. Тестирование когнитивных задач проводилось через 30 минут после введения животным нефирацетама, при этом пик концентрации достигался в течение 2 часов после введения [1].
Для того чтобы вызвать экспериментальную травму мозга, использовалось устройство для перкуссии жидкости, состоящее из цилиндрического резервуара длиной 60 см и диаметром 4,5 см. Хирургическая подготовка началась с установки на один конец устройства насадки Luer-Lock, которая затем имплантировалась поверх открытой твердой мозговой оболочки крыс. Крысы были подготовлены к операции за 24 часа до операции и им вводили 54 мг/кг пентобарбитала натрия в целях анестезии. Была проведена краниэктомия и были сделаны отверстия для фрез, чтобы модифицированная ступица шприца Luer-Lock могла быть помещена поверх открытой и неповрежденной твердой мозговой оболочки. Шприц был прикреплен к твердой мозговой оболочке с помощью цианоакрилатного клея и стоматологического акрила, и животные были помещены обратно в свои домашние клетки, где за ними внимательно наблюдали в процессе восстановления.
Через 24 часа после завершения подготовительной процедуры животных анестезировали с помощью 4% изофлурана, чтобы сделать надрез на коже головы и обнажить ступицу шприца. Группа испытуемых, отнесенных к одной из экспериментальных групп с травмами, была подключена к устройству для нанесения травмы, чтобы нанести травму. Группы с ложной травмой следовали той же процедуре, однако устройство для нанесения травмы так и не было активировано, поэтому животным не было нанесено никакой травмы. Разрез был закрыт, и животные вернулись в свою домашнюю клетку, где за ними наблюдали и обеспечивали свободный доступ к пище и воде. Основными показателями результата, оцениваемыми после травмы, были правильные реакции и выполнение водного лабиринта Морриса из-за связи между травмой от перкуссии жидкости и различными когнитивными дефицитами и неврологическими нарушениями, такими как арефлексия, ступор и потеря сознания [1].
Во-первых, реакция выпрямления использовалась для измерения посттравматического бессознательного состояния у испытуемых, поскольку это сложный постуральный соматический двигательный рефлекс, который обычно подавляется в течение нескольких минут после индукции черепно-мозговой травмы. Животное помещали на спину, и исследователи регистрировали количество времени, которое требовалось им для того, чтобы выпрямиться после травмы. Кроме того, когнитивные функции измерялись с помощью теста водного лабиринта Морриса. Чистая платформа была помещена в 1 из 4 квадрантов в испытательном аппарате, который затем заполнялся водой до тех пор, пока верхняя часть платформы не была скрыта от крыс. 4 тестовых испытания с 4-минутными интервалами между испытаниями проводились на 11-15 день после индукции черепно-мозговой травмы, чтобы обеспечить восстановление любых остаточных двигательных дефицитов [1].
Испытание началось с помещения крысы в бассейн лицом к стене в одном из четырех стартовых положений: север, юг, восток и запад, и им было дано 120 секунд, чтобы найти скрытую платформу. Компьютерная программа отслеживания использовалась для записи активности животных и количества времени, которое потребовалось им, чтобы достичь платформы. Программа также измеряла расстояние от крысы до платформы во время каждого следа и суммировала расстояние, измеренное с интервалом в 0,2 секунды, чтобы вычислить «кумулятивное расстояние от цели» [1]. Оно определялось как то, насколько близко каждое животное плыло к платформе на протяжении всего испытания. Исследователи упомянули, что они также отслеживали скорость плавания животных во время каждого испытания, чтобы обнаружить наличие каких-либо длительных двигательных нарушений.
2) Оценка классического состояния моргания у кроликов полезна для выводов относительно обучения, памяти и старения, поскольку эти поведенческие и нейробиологические нарушения являются отличительными признаками болезни Альцгеймера. Болезнь Альцгеймера также характеризуется деменцией, связанной с нейропатологической потерей нейронов и накоплением амилоидсодержащих бляшек и нейрофибриллярных клубков. Подобная патология наблюдалась при вскрытии людей старше 35 лет с синдромом Дауна; это состояние называется синдромом Дауна/болезнью Альцгеймера (СД/БА).
Предыдущие исследования показывают, что развитие болезни Альцгеймера коррелирует с нарушением классического условного рефлекса моргания больше, чем с типичными нарушениями, наблюдаемыми при старении. Интересно отметить, что взрослые с синдромом Дауна/синдромом Альцгеймера также плохо справлялись с классическим условным рефлексом моргания, однако взрослые с синдромом Дауна набрали значительно больше баллов, чем люди с синдромом Дауна/синдромом Альцгеймера или болезнью Альцгеймера. Дефициты, наблюдаемые в классическом условном рефлексе моргания, связанные с болезнью Альцгеймера, указывают на потенциальное нарушение функционирования холинергических систем в гиппокампе. При этом болезнь Альцгеймера, по-видимому, влияет на функционирование гиппокампа, нарушая холинергическую иннервацию как корковых, так и гиппокампальных нейронов, что приводит к более длительной скорости приобретения классического условного рефлекса моргания. Для выявления потенциального лечения этих нарушений использовался нефирацетам из-за его предполагаемой способности стимулировать высвобождение различных нейротрансмиттеров, которые могут способствовать нормальному функционированию гиппокампа [2].
Исследователь Диана С. Вудрафф-Пак выдвинула гипотезу, что нефирацетам влияет на классическое условное рефлексообразование, улучшая функционирование пирамидальных клеток гиппокампа. При этом введение ноотропа должно улучшить классическое условное рефлексообразование у животных с неповрежденным гиппокампом, но не вызовет никаких изменений у животных, у которых гиппокамп был удален. В эксперимент 1 были включены 56 новозеландских кроликов-пенсионеров без специфических патогенов (SPF), а в эксперимент 2 были включены еще 11 кроликов SPF с подтвержденными поражениями. Средний возраст кроликов, включенных в эксперимент 1, составил 27,9 месяцев, тогда как средний возраст кроликов, включенных в эксперимент 2, составил 26,8 месяцев. Все животные содержались в стандартных лабораторных условиях и имели свободный доступ к кроличьей пище и водопроводной воде [2].
Для удержания каждого кролика во время классических условий моргания использовалось сдерживающее приспособление. Ретракторы удерживали левый глаз открытым, в то время как съемная платформа, удерживающая нагревательную площадку на месте, была прикреплена к морде животных за ушами. Головная площадка располагалась на расстоянии 1 см от роговицы, а безусловный стимул подавался через воздушный поток. Измерения движения мигательной мембраны (ММ) также собирались с помощью мини-потенциометра крутящего момента, прикрепленного к головной площадке с помощью уровня и провода к нейлоновому шву в ММ. Потенциометр на креплении головы преобразовывал движения ММ в электрические сигналы, которые подвергались аналого-цифровому анализу. Собранные значения анализировались сразу после каждого сеанса обучения и сохранялись для дальнейшего изучения [2].
Кролики были обездвижены, как описано ранее, для проведения как обучающих сессий, так и инъекций лекарств. Для проведения обучающих сессий животным подкожно вводили нефирацетам или растворитель за 15 минут до начала обучения. Затем кроликов помещали в ограничитель, оснащали креплением для головы и помещали в вентилируемую и звукоизолированную экспериментальную камеру, где условный стимул подавался в сочетании с безусловным стимулом – струей воздуха в роговицу.
Условным стимулом был звук длительностью 850 мс, 85 дБ и частотой 1 кГц, за которым следовала подача 100 мс, 3 фунта на квадратный дюйм роговичного воздушного потока, спустя 750 мс. Подача как условного стимула, так и роговичного воздушного потока прекращалась одновременно со случайным интервалом между испытаниями продолжительностью 10-20 секунд. Полный сеанс длился 35-45 минут, а обучение завершалось в течение часа после введения нефирацетама или носителя. Во время испытания условный ответ автоматически подсчитывался компьютерной системой, если движение мигательной перепонки происходило в течение 25-750 мс после начала условного стимула, а также если движение было больше 0,5 мм [2].
В первом проведенном эксперименте 64 кролика с неповрежденным гиппокампом были включены в ежедневные сеансы обучения, которые включали 90 попыток сочетания условного стимула звука и безусловного стимула воздушного потока. Нефирацетам растворяли в стерильном физиологическом растворе и вводили подкожно в дозах 1 мг/кг, 3 мг/кг, 5 мг/кг, 10 мг/кг и 15 мг/кг. Во втором эксперименте в исследование были включены 11 кроликов, у которых гистологически было подтверждено двустороннее хирургическое удаление гиппокампа, в то время как еще 20 кроликов прошли хирургическую процедуру для создания аспирационных поражений в асептических хирургических условиях [2].
Кроликам, перенесшим операцию по созданию аспирационных поражений, давали восстановиться в течение 3 недель, прежде чем все испытуемые проходили дальнейшие испытания на выработку условного рефлекса. Нефирацетам растворяли в стерильном физиологическом растворе и вводили животным в дозах 0 мг/кг, и им давали только физиологический раствор или 10 мг/кг. Испытания на выработку классического условного рефлекса моргания проводились по тем же методам, что и в предыдущих экспериментах. Показатели кроликов, перенесших гиппокампэктомию, сравнивали с показателями кроликов, которым делали инъекцию стерильного физиологического раствора, и кроликов, которым вводили дозу нефирацетама 10 мг/кг [2].
1) Результаты оценки рефлекса выпрямления, проведенной исследовательской группой ДеФорда и др., показали, что животные, подвергшиеся индукции черепно-мозговой травмы, испытали значительно более длительное подавление правильной реакции по сравнению с группами, подвергшимися ложной травме. Эти результаты свидетельствуют о том, что тяжесть травм среди травмированных групп была эквивалентной.
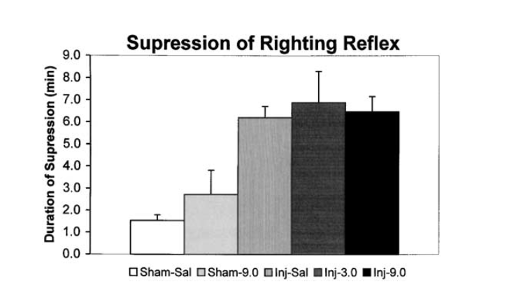
Рисунок 1: Влияние черепно-мозговой травмы на продолжительность подавления рефлекса выпрямления в каждой экспериментальной группе лечения.
Что касается результатов теста с водным лабиринтом Морриса, то короткая латентность цели была показателем лучшей производительности и ответа на лечение. Сообщалось, что у травмированных животных, которым вводили как 3 мг/кг, так и 9 мг/кг нефирацетама, латентность цели была значительно короче по сравнению с травмированными животными, которым вводили физиологический раствор. Кроме того, не было никаких существенных различий между группами ложного воздействия или контрольными группами без травм и теми, кто получал 9 мг/кг нефирацетама. Эти результаты показывают, что лечение ноотропом может восстановить латентность цели до уровня, аналогичного исходному [1].
Рисунок 2: Влияние черепно-мозговой травмы на латентность достижения цели в экспериментальных группах, прошедших тестирование в водном лабиринте Морриса на 11–15-й день после травмы.
Как уже упоминалось ранее, исследователи использовали программу отслеживания активности для измерения «кумулятивного расстояния от цели», определяемого как расстояние, которое каждое животное находилось от платформы во время испытания. Более низкие баллы указывают на лучшую производительность. Подобно полученным результатам задержек цели, кумулятивное расстояние травмированных животных, которым вводили как 3 мг/кг, так и 9 мг/кг нефирацетама, было значительно ниже по сравнению с травмированными животными, которым вводили физиологический раствор. Кроме того, не было никаких существенных различий между баллами ложных групп и тех, кому вводили 9 мг/кг нефирацетама. Эти результаты указывают на то, что лечение ноотропом может восстанавливать кумулятивные расстояния от платформы до уровня, аналогичного исходному [1].
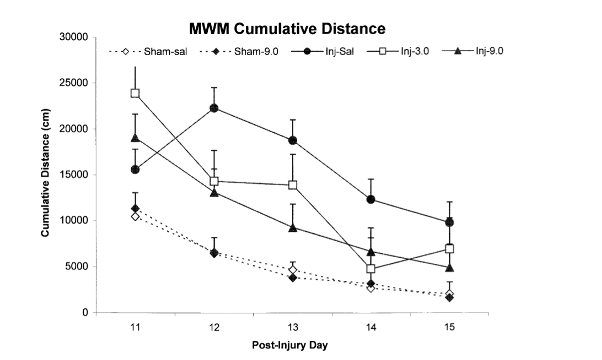
Рисунок 3: Влияние черепно-мозговой травмы на совокупное расстояние от платформы, зафиксированное в экспериментальных группах, прошедших тест с водным лабиринтом Морриса.
Что касается скорости плавания животных, однофакторный ANOVA показал, что не было никаких существенных различий в скорости плавания, показанных между экспериментальными группами. Эти результаты предположили для исследователей, что измеренные различия в задержке цели и кумулятивном расстоянии от платформы не были связаны с двигательными нарушениями, вызванными травмой, или побочными эффектами лечения лекарствами [1].
2) Исследование, проведенное Вудрафф-Пак, изучало, как введение нефирацетама влияет на классическое условное моргание у кроликов с неповрежденным гиппокампом и у кроликов с двусторонне удаленным гиппокампом. Было проведено два эксперимента, в первом из которых 64 интактных кролика получали различные дозы ноотропа, включая 1, 3, 5, 10 и 15 мг/кг. Во втором эксперименте участвовали 11 кроликов с удаленным гиппокампом и 20 кроликов, перенесших операцию по созданию аспирационных поражений на гиппокампе. Затем их лечили либо стерильным физиологическим раствором, либо 10 мг/кг нефирацетама [2].
Первоначальные результаты первого эксперимента показали, что у кроликов, которым вводили нефирацетам, наблюдалось значительное улучшение количества времени, необходимого для изучения связи между доставленными стимулами, по сравнению с кроликами, которым вводили только растворитель. Улучшение времени обучения свидетельствует об улучшении когнитивных функций; доза, которая оказалась наиболее эффективной, составила 10 мг/кг нефирацетама. Также было обнаружено, что амплитуда безусловной реакции была наиболее существенно затронута дозами нефирацетама 3 мг/кг по сравнению с животными, которым вводили растворитель. Важно отметить, что у кроликов, которым вводили 10 мг/кг ноотропа, средняя амплитуда безусловной реакции была сопоставима с кроликами, которым вводили растворитель. Амплитуда в группе лечения 10 мг/кг была зарегистрирована как 4,38 мм, в то время как амплитуда в группе лечения растворителем была зарегистрирована как 5,00 мм [2]. Это произошло несмотря на предыдущие результаты, согласно которым доза нефирацетама 10 мг/кг смягчает дефицит обучения и улучшает когнитивные функции.
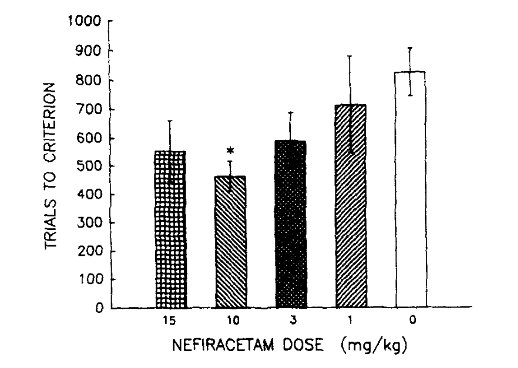
Рисунок 4: Количество попыток, которое потребовалось старшим кроликам, включенным в каждую экспериментальную группу лечения, чтобы усвоить связь между безусловным и условным стимулами.
Результаты второго эксперимента, проведенного с участием кроликов с удаленным гиппокампом, показали, что у здоровых старых кроликов, которым вводили 10 мг/кг нефирацетама, наблюдалась значительно лучшая скорость обучения по сравнению с теми, кому давали плацебо. При этом лечение 10 мг/кг нефирацетама не вызвало никаких существенных эффектов на обучение у кроликов с удаленным гиппокампом. Эти результаты подтверждают гипотезу, разработанную Вудраффом-Паком, о том, что лечение ноотропом не влияет на когнитивные функции у кроликов, у которых нет гиппокампа, но улучшает их у кроликов, у которых гиппокамп остался неповрежденным. Это указывает на то, что механизм действия, который обуславливает преимущества нефирацетама, связан с пирамидальными клетками гиппокампа [2].
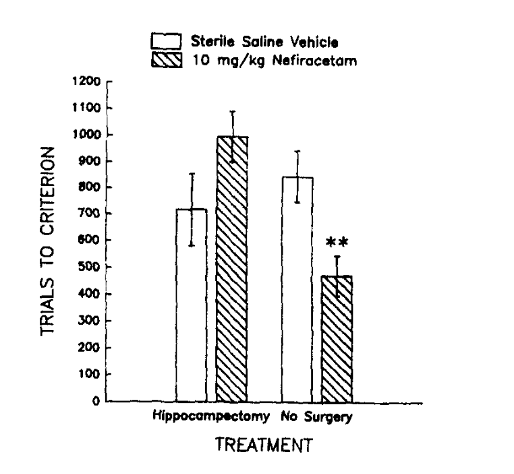
Рисунок 5: Количество попыток, которое потребовалось гиппокампэктомированным и интактным кроликам из каждой экспериментальной группы лечения, чтобы усвоить связь между безусловным и условным стимулами.
Общие результаты экспериментов 1 и 2 показывают, что 10 мг/кг нефирацетама имели потенциал для сокращения количества попыток, необходимых старым кроликам для изучения связи между безусловным и условным стимулом, подаваемым во время тестирования классического условного рефлекса моргания. Кроме того, эта доза была эффективна только для кроликов, у которых не был удален гиппокамп, что указывает на то, что ноотроп ослабляет классическое условное рефлекс моргания, воздействуя на клетки гиппокампа. В частности, пирамидальные нейроны CA1 больше всего страдают от старения, при этом взрослые животные теряют 15-20% этих клеток [2].
При этом было показано, что нефирацетам обладает потенциалом защиты мембранной дисфункции в нейронах гиппокампа CA1 после эпизодов недостатка глюкозы и кислорода. Кроме того, нефирацетам может бороться с дефицитом обучения у пожилых животных, усиливая функцию пирамидальных нейронов гиппокампа CA1, которые все еще сохраняются. Важно отметить, что результаты, касающиеся эффектов нефирацетама, были подтверждены исследованиями, сосредоточенными на другой добавке, усиливающей когнитивные функции, нимодипине. Результаты показывают, что улучшение дефицита классического условного рефлекса моргания у пожилых кроликов связано с облегчением функционирования пирамидальных нейронов гиппокампа [2].
[1] DeFord SM, Wilson MS, Gibson CJ, Baranova A, Hamm RJ. Nefiracetam improves Morris water maze performance following traumatic brain injury in rats. Pharmacol Biochem Behav. 2001 Jul-Aug;69(3-4):611-6. doi: 10.1016/s0091-3057(01)00559-7. PMID: 11509223.
[2] Woodruff-Pak DS. Nefiracetam ameliorates learning deficits in older rabbits and may act via the hippocampus. Behav Brain Res. 1997 Feb;83(1-2):179-84. doi: 10.1016/s0166-4328(97)86065-2. PMID: 9062680.
© 2026 АнкеБио. Magazin nootropov Biohaker
Казахстан, Алматы, ул. Байзакова, 280
Все права защищены.
+7 (977) 520-97-00
Email: ankebiorus@gmail.com
Вы нашли для себя препарат, который хотите
приобрести по низкой цене и с гарантией
безопасной доставки?
Звоните нам
Мы Вам обязательно ответим!



