
График работы:
E-mail:
Бесплатная консультация
Aniracetam (Анирацетам) (1-анизоил-2-пирролидинон) является агонистическим соединением AMPA, которое показало потенциал для регулирования рецептора α-амино-3-гидрокси-5-метил-4-изоксазол пропионата. Механизм действия, лежащий в основе регулирования рецепторов AMPA, включал ингибирование внутри рецептора с целью десенсибилизации субъединицы GluR2 путем снижения уровня проницаемости Ca2+. Текущие исследования, касающиеся анирацетама, фокусируются на способности соединения вызывать эмоциональные расстройства, нарушения цикла сна и когнитивные функции, поскольку это связано с такими состояниями коры головного мозга, как инсульты, надъядерный паралич, болезнь Альцгеймера и болезнь Паркинсона [1][2].
1) Регуляция рецептора AMPA посредством лечения анирацетамом приводит к улучшению неврологического дефицита и снижению процента инфарктов благодаря нейропротекторным и противовоспалительным свойствам этого соединения.
2) Было показано, что лечение анирацетамом in vitro снижает уровень апоптотических клеток, а также улучшает клеточный окислительный стресс за счет снижения количества присутствующих активных форм кислорода.
1) Исследовательская группа Шармы и соавторов оценила влияние лечения анирацетамом на активность рецепторов AMPA в острой и подострой фазе неврологического повреждения после инсульта. Для целей данного исследования использовались самцы крыс Wistar в возрасте 14–16 недель весом от 240 до 280 граммов. Испытуемые содержались по 4 крысы в клетке в стандартных лабораторных условиях с неограниченным доступом к воде и сухим гранулам. Животным давали возможность акклиматизироваться в течение одной недели, прежде чем их случайным образом распределяли по различным экспериментальным группам лечения. Важно отметить, что в данном исследовании использовались только самцы крыс, чтобы не допустить искажения полученных данных нейропротекторными эффектами эстрогена. Животных анестезировали 5% изофлураном, в то время как церебральная ишемия была вызвана с помощью интралюминальной техники, предназначенной для окклюзии средней мозговой артерии, которая была удалена через 90 минут [1].
Исследовательская группа отслеживала несколько объективных показателей в ходе данного исследования. Во-первых, влияние рецептора AMPA, перампанела, на различные нейроповеденческие параметры после введения дозы 1,5 мг/кг через 10–15 минут после реперфузии. Во-вторых, изучалось влияние перампанела на различные нейроповеденческие параметры после введения дозы 1,5 мг/кг через 10–15 минут после реперфузии в течение 3 дней. В-третьих, изучалось влияние перампанела на различные нейроповеденческие параметры после введения дозы 1,5 мг/кг через 10–15 минут после реперфузии в течение 5 дней. В рамках задачи 4 был введен анирацетам и оценен эффект дозы 50 мг/кг агониста рецептора AMPA при введении через 3 дня после реперфузии, в то время как в рамках задачи 5 оценивался эффект дозы 50 мг/кг анирацетама, введенной через 5 дней после реперфузии. Для цели 6 исследовательская группа определила время, в которое перампанел и анирацетам были наиболее эффективны с точки зрения снижения процента инфаркта и двигательной некоординации. Эти временные точки использовались для вторичных экспериментальных целей [1].
Следующая часть исследования включала нейроповеденческую оценку на основе оценки неврологического дефицита, теста с вращающимся стержнем и теста на силу захвата. После того, как был вызван инсульт средней мозговой артерии, неврологический дефицит, испытываемый испытуемыми, оценивался по 5-балльной шкале; оценка 0 указывала на отсутствие неврологического дефицита; оценка 1 включала невозможность полностью выпрямить правую переднюю лапу; оценка 2 обозначалась кружением вправо; оценка 3 определялась падением вправо; а оценка 4 включала отсутствие спонтанной ходьбы и общее снижение уровня сознания [1].
С другой стороны, тест с вращающимся стержнем исследовал двигательную координацию крыс как до, так и после индукции инсульта средней мозговой артерии. Животным была разрешена одна сессия тренировок с вращающимся аппаратом с вращающимся стержнем; вращающийся стержень двигался с постоянной скоростью 8 миль в час, и сессия тренировок продолжалась до тех пор, пока животные не могли оставаться на вращающемся стержне в течение 60 последовательных секунд. После обучения животные получили базовое испытание, в котором скорость вращающегося стержня увеличивалась с 4 миль в час до 40 миль в час в течение 5 минут, то же самое обследование проводилось на 7-й день после окклюзии [1].
Последним нейроповеденческим тестом был тест на силу захвата, который проводился как до, так и после реперфузионного повреждения. Тест включал в себя устройство, поднятое на 40 см от поверхности и состоящее из прочного натянутого между двумя вертикальными опорными балками 50 см в длину. Испытуемый помещался в среднюю точку между двумя опорными поверхностями и оценивался по 6-балльной шкале; оценка 0 означает, что животное упало с веревки; оценка 1 включает висение на веревке двумя передними лапами; оценка 2 характеризуется попыткой снова забраться на веревку; оценка 3 определяется как висение на веревке двумя передними лапами плюс одна или обе задние лапы; оценка 4 включает висение на веревке всеми лапами и хвостом, обмотанными вокруг веревки; и оценка 5 означает, что животное смогло освободиться от устройства, переместившись к одной из систем поддержки [1].
2) Исследовательская группа Габриэля и соавторов изучила способность анирацетама защищать культивируемые астроциты от ишемического повреждения. После получения необходимых химикатов и материалов астроциты были выделены из однодневных крыс Wistar и культивированы в соответствии со стандартной процедурой. Клетки были высеяны в соответствии с тем, предназначались ли они для биолюминесцентного исследования, флуоресцентного исследования, анализа активности каспазы-3, анализа ОТ-ПЦР или окрашивания Hoechst 33342 [2]. Образцы астроцитов культивировались в среде, состоящей из 20% FBS, которую заменяли через 4 дня средой из 10% FBS; через 1 неделю весь объем культуральной среды меняли дважды в неделю, при этом культуры встряхивали в течение ночи для удаления любых загрязняющих неастроглиальных клеток. Для идентификации астроцитов культуры были иммуногистохимически окрашены для анализа GFAP; исследователи отметили, что все эксперименты проводились на 21-дневных культурах [2].
Экспериментальные процедуры начинались с культивирования нормоксических астроцитов в среде DMEM с 10% FBS и 5,5 мМ глюкозы, после чего культуры помещались в среду DMEM с 3% O2/5% CO2/92% N2 на 24 часа для имитации ишемии in vivo. Анирацетам растворялся в ДМСО в начальной концентрации 10 мМ; затем клеточные культуры обрабатывались различными дозами соединения в диапазоне от 1, 10 и 100 мкМ в течение 24 часов в нормоксии и еще 24 часа имитации ишемии. Ингибитор киназы, PD98059, добавлялся в культуральную среду в концентрации 50 мМ, после чего к культуре добавлялся вортманнин в концентрации 0,1 мМ [2].
Затем апоптоз оценивали путем окрашивания ядер клеток Hoechst 3342 для визуализации фрагментированной и конденсированной ДНК. Астроциты промывали PBS и культивировали в течение 10 минут с 4% параформальдегидом; после второй промывки PBS образцы сначала дегидратировали в 70% этаноле, а затем снова дегидратировали в абсолютном этаноле. Клетки окрашивали Hoechst 33342 и промывали PBS в последний раз, что позволяло провести анализ ядер клеток с помощью флуоресцентной визуализации с использованием рабочей станции MiraCal Pro III и инвертированного микроскопа Eclipse TE200. Исследовали 6 случайно выбранных областей из каждой пластины для культивирования клеток с целью определения количества апоптотических ядер [2]. Каждая область содержала приблизительно 200 клеток, и результаты основывались на расчете процента апоптотических клеток относительно общего количества клеток, обнаруженных в области.
Концентрации АТФ и креатинфосфата были исследованы с помощью высокоспецифичного биолюминесцентного анализа люциферина-люциферазы светлячков. Превращение ПЦр в АТФ катализировалось добавлением креатинкиназы с последующим измерением концентрации АТФ с помощью количественной биолюминесценции. Эти процедуры включают разрушение клеток, обнаруженных в образцах объемом 100 мкл, и добавление их в 5% раствор ТХУ с 2 мМ ЭДТА с последующим быстрым встряхиванием и инкубацией. Затем образцы были разбавлены в 20 раз буфером Трис-ацетата с pH 7,75. Затем 10 мкл концентрации АТФ было добавлено в каждую клеточную пластину в качестве внутреннего стандарта, что позволило исследователям подсчитать общее количество присутствующего АТФ [2].
Исследователи определили ферментативную активность соединения каспазы-3 с помощью колориметрического анализа каспазы-3, который использует высвобождение флуорохрома p-нитроанилина (p-NA) в сочетании с пептидным субстратом, специфичным для каспазы-3. Эти параметры были выбраны исследовательской группой с учетом того, что расщепление пептида посредством активной каспазы-3, как было показано, высвобождает хромофор p-NA, который можно количественно определить с помощью колориметрического планшетного ридера 405 нМ. Основной механизм количественной оценки включал лизирование культивируемых астроцитов и центрифугирование клеточных экстрактов для устранения любого дебриса [2]. Затем 20 мкл клеточных экстрактов инкубировали с субстратом, и уровень активности каспазы-3 рассчитывали как пропорцию интенсивности цветной реакции к контрольным значениям.
Кроме того, было изучено превращение 2′,7′-DCF-DA во флуоресцентный DC, опосредованное присутствием активных форм кислорода, с целью определения уровней клеточного окислительного стресса. Культивируемые астроциты подвергались воздействию 100 мкМ DCF-DA и инкубировались в течение 50 минут, после чего их трижды промывали HBSS, в то время как для количественной оценки флуоресценции DCF использовался микропланшетный ридер Fluoroscan. Краситель, используемый для этой части исследования, возбуждался при 485 нм, а излучение фильтровалось с использованием барьера, установленного на 538 нм; исследовательская группа определяла продукцию активных форм кислорода в процентах от контрольных клеток.
Наконец, был завершен анализ ОТ-ПЦР с целью определения экспрессии генов c-fos и c-jun в культивируемых образцах астроцитов крыс после воздействия ишемических условий в течение 24 часов и обработки различными дозами анирацетама. Общая РНК была выделена из астроцитов и культивирована на 100-миллиметровых пластинах для культивирования клеток. РНК была осаждена и очищена с последующим растворением в стерильной воде без нуклеазы и сохранена для дальнейшего использования. Выделенная РНК была количественно определена, и реакция обратной транскрипции была проведена с использованием 100 нг общей РНК [2].
Амплификация экспрессии генов проводилась в течение 30 циклов для праймеров c-fos, c-jun и бета-актина; эти праймеры, используемые в анализе ПЦР, соответствовали определенной последовательности ДНК для каждого из генов. ОТ и ПЦР проводились с помощью термоциклера Termoblock, и 10 мкл амплифицированных продуктов ПЦР окрашивались бромистым этидием в 1% буфере TAE и анализировались денситометрией. Все эксперименты были выполнены трижды, и результаты были количественно оценены с помощью программного обеспечения Image Pro Plus, с последующей нормализацией в соответствии с экспрессией гена бета-актина [2].
1) Первоначально было подтверждено, что окклюзия средней мозговой артерии привела к значительному увеличению неврологического дефицита и снижению баллов по тестам на силу захвата и вращающийся стержень. Результаты первых трех задач, оценивающих эффективность распространенного агониста рецепторов AMPA, перампанела, показали, что при введении соединения через 3 и 5 дней после индукции инсульта средней мозговой артерии неврологический дефицит был значительно снижен. Хотя введение в оба эти периода времени снижало процент инфаркта, было обнаружено, что через 5 дней после окклюзии наблюдалось максимальное восстановление. Те же результаты были получены в тестах на вращающийся стержень и силу захвата, где максимальное восстановление наблюдалось на 5-й день после окклюзии. Основное различие между тестами на вращающийся стержень и силу захвата и оценкой неврологического дефицита заключалось в том, что агонист рецепторов AMPA вводился на 1-й день после окклюзии, а также на 3-й и 5-й дни [1].
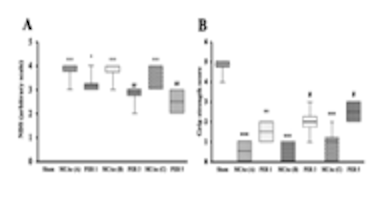
Рисунок 1: Изменения A) показателя неврологического дефицита, B) показателя силы захвата, C) времени, проведенного на вращающемся стержневом аппарате, в ответ на лечение парампанелом после инсульта средней мозговой артерии.
Что касается задач 4 и 5, изначально было подтверждено, что окклюзия средней мозговой артерии привела к увеличению баллов неврологического дефицита, а также к снижению баллов по тесту на вращающийся стержень и силу захвата. Результаты этой части исследования показали, что лечение анирацетамом как на 2-й, так и на 5-й день после ишемического реперфузионного повреждения имело потенциал для получения максимальной защиты от неврологического дефицита. Что касается тестирования силы захвата, лечение анирацетамом, вводимое как на 3-й, так и на 5-й день после ишемического реперфузионного повреждения, значительно улучшило баллы силы захвата. Однако важно отметить, что введение соединения на 5-й день вызвало более сильные эффекты, чем при введении на 3-й день. Наконец, результаты теста на вращающийся стержень показали, что при введении анирацетама животные проводили значительно больше времени в аппарате с вращающимся вращающимся стержнем. Было обнаружено, что улучшение производительности и координации движений было наиболее значительным, когда анирацетам вводился через 5 дней после ишемического реперфузионного повреждения [1].

Рисунок 2: Изменения A) показателя неврологического дефицита, B) показателя силы захвата, C) времени, проведенного на вращающемся стержневом аппарате, в ответ на лечение анирацетамом после инсульта средней мозговой артерии.
Наконец, цель 6 изучала эффекты последовательного лечения перампанелом и анирацетамом на неврологический дефицит, мышечную силу, оцениваемую с помощью теста на силу захвата, и координацию движений, оцениваемую с помощью теста с вращающимся стержнем. После индукции ишемии средней мозговой артерии были выявлены значительные неврологические нарушения, которые в конечном итоге были устранены после последовательного лечения перампанелом и анирацетамом. Аналогичные результаты были получены для теста на силу захвата и теста с вращающимся стержнем, что указывает на улучшение мышечной силы и координации движений соответственно после последовательного лечения перампанелом и анирацетамом [1].

Рисунок 3: Изменения A) показателя неврологического дефицита, B) показателя силы захвата, C) времени, проведенного на вращающемся стержневом аппарате, в ответ на последовательное лечение парампанелом и анирацетамом после инсульта средней мозговой артерии.
2) При изучении результатов окрашивания Hoechst 33342 с целью определения влияния анирацетама на апоптоз. После того, как клеточные культуры подвергались воздействию 24 часов имитируемых ишемических условий, количество апоптотических ядер значительно увеличивалось. При обработке культур различными дозами анирацетама количество апоптотических ядер заметно снижалось, причем доза ноотропа 100 мкМ оказывала наиболее сильное действие против апоптоза, вызванного ишемией. Исследовательская группа также исследовала роль PI 3-киназы и MAP-киназы/MEK в антиапоптотическом механизме анирацетама. При этом 0,1 мкМ ингибитора PI 3-киназы, вортманнина, и 50 мкМ ингибитора MEK, PD98059, были использованы для того, чтобы помочь исследователям в дальнейшем определении механизма, лежащего в основе предотвращения гибели клеток [2].
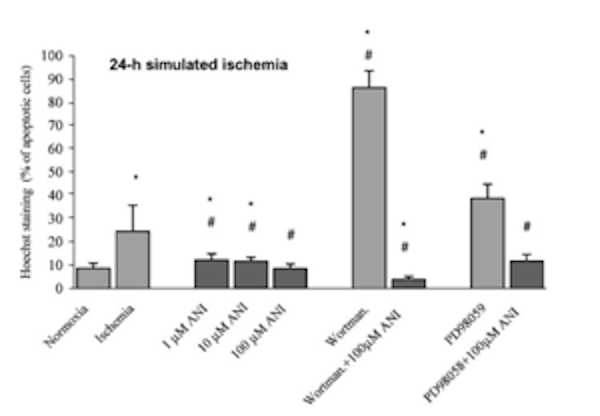
Рисунок 4: Изменения процентного содержания апоптотических клеток, измеренные с помощью Hoechst 33342, в ответ на обработку 1, 10 или 100 мкМ анирацетама.
Клеточные культуры, обработанные вортманнином и PD98059 во время воздействия ишемических условий, привели к гибели 86 +/- 7,3% и 38,7 +/- 6% клеток соответственно. При введении 100 мкМ анирацетама в сочетании с вортманнином или PD98059 количество апоптотических ядер снизилось до 3,9 + 1,3% и 11,8 + 2,9% соответственно. Эти результаты позволили исследовательской группе сделать вывод о том, что анирацетам оказывает значительное влияние на пути MEK и PI 3-киназы [2]. Доза ноотропа 100 мкМ также имела потенциал для предотвращения процесса апоптоза при воздействии ишемических условий.
После воздействия на культуры астроцитарных клеток имитируемых ишемических условий, клетки обрабатывали дозами анирацетама в диапазоне от 1, 10 и 100 мкМ для того, чтобы исследователь мог изучить влияние соединения на уровень креатинфосфата и АТФ. Было обнаружено, что обработка анирацетамом значительно увеличивает концентрацию АТФ, однако не было обнаружено никаких заметных эффектов на уровни креатинфосфата в образцах [2]. Кроме того, при наблюдении за влиянием ноотропа на активность каспазы-3, 1 мкМ анирацетама не вызывал никаких существенных эффектов на ферментативную активность, в то время как дозы 10 и 100 мкМ значительно снижали активность каспазы-3 в образцах, подвергнутых ишемическим условиям [2].
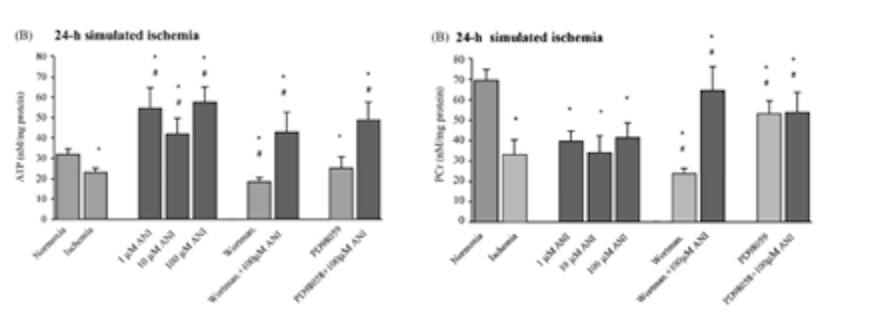
Рисунок 5: Изменения внутриклеточного АТФ и ФКр в ответ на обработку 1, 10 или 100 мкМ анирацетама.
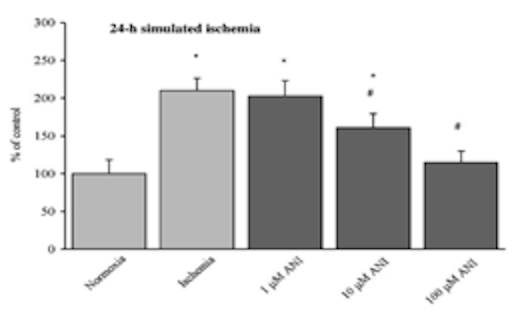
Рисунок 6: Изменения ферментативной активности каспазы-3 в ответ на обработку 1, 10 или 100 мкМ анирацетама.
Исследователи также оценили влияние анирацетама на клеточный окислительный стресс, отмеченный наличием активных форм кислорода. Было обнаружено, что анирацетам вызывает резкое снижение флуоресценции DCF в образцах астроцитов, что указывает на заметное снижение продукции активных форм кислорода. Астроциты также подвергались ишемическим условиям в сочетании с ингибитором киназы PD98059, что приводило к увеличению продукции активных форм кислорода. Этот эффект ослаблялся при использовании 100 мкМ анирацетама для обработки культур астроцитов [2].
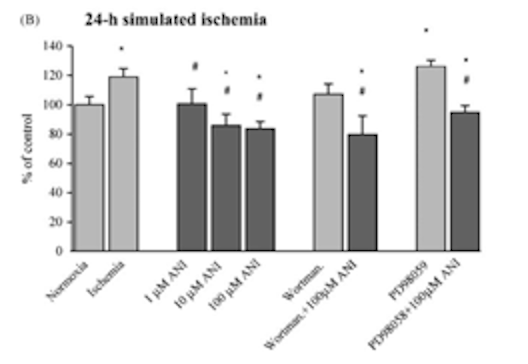
Рисунок 7: Изменения в продукции активных форм кислорода в ответ на лечение 1, 10 или 100 мкМ анирацетама.
Аналогичные результаты были получены при изучении эффектов соединения на экспрессию генов c-fos и c-jun. 24-часовое воздействие имитируемых ишемических условий привело к увеличению экспрессии обоих генов, а лечение 10 и 100 мкМ анирацетама в течение 24 часов, как было обнаружено, значительно снизило экспрессию генов. Важно отметить, что доза ноотропа 1 мкМ не оказала существенного влияния на экспрессию генов [2].
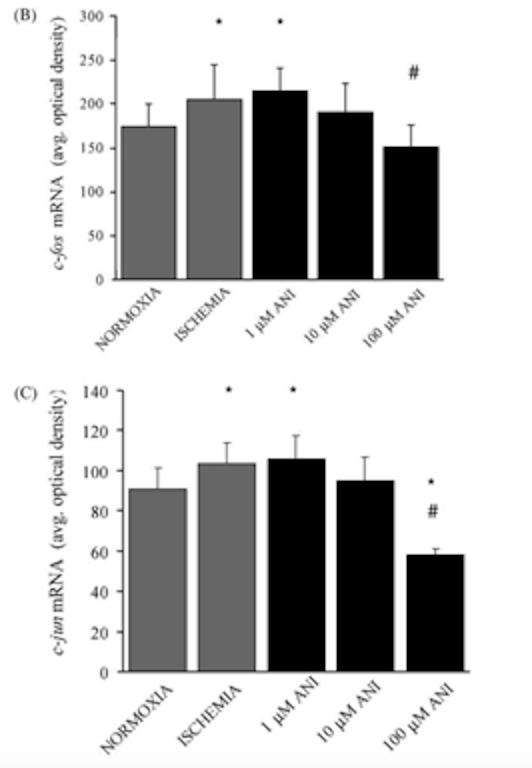
Рисунок 8: Изменения экспрессии генов c-fos и c-jun в ответ на лечение 1, 10 или 100 мкМ анирацетама.
Анирацетам – жирорастворимая молекула, принадлежащая к семейству рацетамов. Анирацетам тесно связан с популярным ноотропным соединением пирацетамом, но часто считается более эффективным. Различия между двумя соединениями обусловлены его структурными компонентами; анирацетам заменяет аминогруппу пирацетама на метилированную фенильную группу, чтобы повысить растворимость в жире. Подобно многим другим соединениям рацетама, анирацетам изначально метаболизируется через печеночную систему, однако, данные показывают, что он также эффективно метаболизируется через кишечник, даже в состоянии голодания. Соединение наиболее известно своим воздействием на нейротрансмиссию, а также своей способностью модулировать возбуждающие рецепторы AMPA. При этом анирацетам изучается на предмет его способности лечить депрессию и тревожность,
а также нарушения памяти.
Исследования на животных изучали эффекты анирацетама, действующего как модулятор рецептора AMPA. Анирацетам работает как эффективный модулятор рецепторов AMPA посредством своего механизма связывания с неактивным участком рецептора. Затем соединение аллостерически модифицирует участок связывания, что приводит к снижению скорости десенсибилизации при взаимодействии возбуждающих стимулов, таких как глутамат, с рецептором. Важно отметить регуляцию рецепторов AMPA, поскольку они управляют большей частью возбуждающей передачи в мозге. Это приводит к ускоренной передаче аминокислот и улучшению навыков извлечения памяти.
Анирацетам также показал себя многообещающим в регуляции действия каинатных рецепторов, соединений, которые тесно связаны с действием глутаминэргической передачи и активностью рецептора AMPA. Каинатные рецепторы собираются из 5 различных единиц, связанных с глутаматом, GluK1-5. Исследователи Лоури и др. изучали, как удаление одной из этих субъединиц влияет на память у мышей. Первоначально мыши с нокаутом GluK4 были подвергнуты водному лабиринту памяти и продемонстрировали резкие нарушения приобретения памяти и припоминания.
У этих мышей также наблюдались признаки гиперактивности и нарушения преимпульсного торможения – два характерных симптома шизофрении и биполярного расстройства. При сравнении мышей с нокаутом GluK4 с контрольной группой дикого типа были замечены огромные различия в памяти, что привело исследователей к мысли, что GluK4 также может играть роль в регуляции психиатрических расстройств. Результаты показали, что водный лабиринт, которому подвергались мыши, в значительной степени зависит от функционирования гиппокампа. Правильное функционирование гиппокампа также связано с шизофренией и потерей памяти.
Данные также свидетельствуют о том, что и шизофрения, и биполярное расстройство связаны с глутаминэргической передачей в мозге, в частности с активностью GluK4. Это позволило исследователям сделать вывод о том, что нейропсихиатрические расстройства и нейродегенерация, такие как потеря памяти, потенциально могут быть вызваны неправильным функционированием Glu4K. Однако необходимо провести более обширные исследования, чтобы полностью понять пути передачи в мозге, а также то, как анирацетам может исправить эти проблемы, действуя как положительный модулятор рецепторов каината и AMPA.
Анирацетам также продемонстрировал потенциал в повышении уровней холинергической и дофаминергической передачи. Дофамин и ацетилхолин (АХ) – два нейротрансмиттера, которые необходимы для лечения тревожности и депрессии, а также для улучшения сохранения памяти и обучения. Исследования на животных показали, что анирацетам способен усиливать сигнализацию никотиновых рецепторов посредством взаимодействия с белками. Кроме того, при введении доз анирацетама 50 мг/кг у крыс наблюдалось снижение скорости оборота дофамина, что приводило к улучшению настроения и памяти.
Благодаря своей способности регулировать рецепторы AMPA и каинат, введение анирацетама улучшает нейропротекцию и потерю памяти. Знаменательное исследование, проведенное Cumin et. Al, вызвало нарушения памяти и обучения с помощью холинергических антагонистов, ишемии и электросудорожного шока. Результаты этого исследования показали, что дозы приблизительно 1,5 г могут обратить нарушения вспять и защитить от новой травмы. Кроме того, два различных исследования сообщили, что регулирование рецептора AMPA может привести к увеличению нейротрофического фактора мозга (BDNF), соединения, широко известного своей способностью улучшать нейронную пластичность. Последующий протокол стимуляции привел к устойчивому повышению уровней BDNF в течение 5 дней.
Дополнительные исследования отметили способность анирацетама влиять на нейротрансмиттеры, такие как дофамин, серотонин и ACh. При введении доз анирацетама, варьирующихся от 10 до 100 мг/кг, исследователи отметили, что тревожность резко снижалась из-за многочисленных взаимодействий между соединением и системами нейротрансмиссии. Известное исследование, проведенное Накамурой и др., подчеркнуло, как усиление дофаминергической сигнализации с помощью анирацетама было эффективным для оказания антидепрессивного эффекта у подопытных крыс в дозах 100 мг/кг. Важно отметить, что метаболит анирацетама, галоперидол и мекамиламин вызывали те же самые эффекты.
[1] Sharma H, Reeta KH, Sharma U, Suri V, Singh S. AMPA receptor modulation through sequential treatment with perampanel and aniracetam mitigates post-stroke damage in an experimental model of ischemic stroke. Naunyn Schmiedebergs Arch Pharmacol. 2023 Dec;396(12):3529-3545. doi: 10.1007/s00210-023-02544-z. Epub 2023 May 25. PMID: 37231168.
[2] Gabryel B, Adamczyk J, Huzarska M, Pudełko A, Trzeciak HI. Aniracetam attenuates apoptosis of astrocytes subjected to simulated ischemia in vitro. Neurotoxicology. 2002 Sep;23(3):385-95. doi: 10.1016/s0161-813x(02)00084-0. PMID: 12387365.
© 2026 АнкеБио. Magazin nootropov Biohaker
Казахстан, Алматы, ул. Байзакова, 280
Все права защищены.
+7 (977) 520-97-00
Email: ankebiorus@gmail.com
Вы нашли для себя препарат, который хотите
приобрести по низкой цене и с гарантией
безопасной доставки?
Звоните нам
Мы Вам обязательно ответим!



